Доказано, что при депрессии нарушаются многие биологические ритмы. В частности, происходит снижение амплитуды выброса кортизола, тиреостимулирующего гормона, мелатонина, числа сердечных сокращений, температуры тела, серотонина и др. (Wehr Т. А., 1977; Kripke D.F. et al., 1978; Schultz H., Lung R., 1983; Duncan W.C., 1996). Заинтересованность серотонинергических механизмов в хронобиологическом механизме развития депрессии косвенно подтверждают высокая концентрация серотонина и высокая плотность 5-НТ2с-рецепторов в супрахиазматическом ядре (СХЯ).
Нарушение цикла сон-бодрствование занимает одно из ведущих мест при депрессии и связано с определенной поломкой центральных механизмов, регулирующих процессы психофизиологической ритмики в организме (Wehr Т. A., Goodwin F. К., 1983). Современные представления о системе синхронизации циркадианных ритмов с циклом сон-бодрствование представлены на рис.З. Кроме типичной циркадианной организации психопатологической симптоматики депрессии с ухудшением состояния утром и улучшением к вечеру, сами фазовые колебания, по-видимому, являются расстроенным инфрадианным ритмом. В частности, это подтверждается существованием давно подмеченной сезонности аффективных нарушений (Rosenthal N. et al., 1983; Lacoste R., Wirz-Justice A., 1989, Magnusson A., Partonen T., 2005) и особенно явственно обнаруживается при континуальном течении или феномене «быстрой цикличности» (БЦ), доходящей в наиболее злокачественных случаях до собственно суточного ритма (Sitaram N. et al., 1978).
Немецкий исследователь Papousek (1975) в качестве гипотезы, объясняющей развитие депрессии, выдвинул теорию фазового опережения циркадианных ритмов, которая в последующем нашла подтверждение в ряде клинических исследований. В частности, обнаруживается десинхронизация ритма сон-бодрствование и температурного циркадианного ритма, а при полисомнографическом исследовании — укорочение латентного периода ПФ сна (ЛПФ), т.е. смещение ее развития на более раннее время суток. Смещение фазы продукции мелатонина в течение ночи коррелирует с выраженностью депрессии (Emens J. et al., 2009). При депрессии обнаруживается нарушение суточного ритма выработки мелатонина и повышение его концентрации в крови при применении антидепрессантов (Sonetre Е. et al., 1989). Хотя сам мелатонин антидепрессивным действием не обладает, он может улучшать архитектуру сна (Srinivasan V. et al., 2006). Аналогичные изменения происходят в эксперименте у здоровых добровольцев при сенсорноинформационной депривации (Lund R., 1989). На этом основании ряд исследователей выдвинул гипотезу о нарушении у больных депрессией центрального механизма, синхронизирующего циркадианные ритмические процессы, что проявляется в своеобразной невосприимчивости к имеющим важное регулирующее значение факторам, таким как смена дня и ночи (Mendlewicz J., 1982; Wehr Т, Goodwin Е К.,1983). У больных аффективными расстройствами и их родственников были обнаружены специальные генные локусы, ответственные за недостаточность функции биологических часов в организме и повышающие риск развития депрессии (Johansson С. et al., 2003).

Рис. 3. Система синхронизации циркадианных ритмов и хронобиологическая гипотеза депрессии
Примечание.
Циркадианные ритмы млекопитающих регулируются центральными биологическими часами (главный пейсмекер), расположенными в парных СХЯ в переднем гипоталамусе, содержащих около 20000 клеток. Ядро задает эндогенный биологический ритм, близкий к 24 часам, по всему организму. В экспериментальных условиях отдельные нейроны СХЯ сохраняют отчетливый циркадианный ритм электрической активности, который регистрируется в течение нескольких недель, слегка отклоняясь от 24 часов (обычно несколько длиннее) (Welsh Р. К. et al., 1995). Этот ритм все время подвергается коррекции (синхронизируется) различными факторами внешней среды (zeitgebers), преимущественно сменой дня и ночи. Фотосигналы передаются в мозг посредством 2 зрительных путей: ретиногипоталамического тракта (РГТ), который осуществляет прямую связь между сетчаткой и СХЯ, и непрямого пути от сетчатки через оптический тракт к боковому коленчатому комплексу, от которого через геникулогипоталамичсский тракт (ПТ) осуществляется связь с СХЯ. Несветовое модулирующее влияние на СХЯ осуществляется из серотонинового медианного ядра шва также двумя путями: прямым восходящим пучком к СХЯ и опосредованным влиянием через интергеникулярную прослойку. Через паравентрикулярное ядро (ПВЯ) возбуждение СХЯ стимулирует выработку гормона мелатонина в эпифизе. ПВЯ также содержит автономные нейроны, передающие сигналы о времени суток разным органам и нейронам, секретирующим кортикотропин-релизинг гормон (CRH), котциркадианныеимпульсы, поступающие из СХЯ, трансформируется в гормональные и вегетативные стимулы для порганов. Такаятрансформация сигнала происходит преимущественно в ПВЯ. Мелатонин сам по себе не определяет циркадианный ритм, а оказывает модулирующее влияние на него, смещая фазу ритма (ее начало и длительность) через воздействие на МТ, рецепторы в СХЯ (Weil Z. М. et al., 2006) и устанавливая чувствительность эндогенных часов к свету, т.е. фактически играет роль их маятника. Высвобождение мелатонина начинается ранним вечером и является наиболее надежным биологическим маркером периодичности циркадианных ритмов. Стимуляция МТ2 рецепторов играет ингибирующую роль в СХЯ (их блокада в эксперименте у животных вызывает депрессивноподобное поведение и снижает амплитуду ритмов). Серотониновое влияние на СХЯ играет модулирующую ограничительную роль. В СХЯ обильно присутствуют 5-НТ2С рецепторы, которые, как известно, принимают участие в регуляции медленноволновой фазы сна (их блокада усиливает дельта-волновую активность, а стимуляция вызывает светоподобный эффект на циркадианные ритмы) (Kennaway D.J., Moyer R. W., 1998).
Нарушения разных эндогенных ритмов при депрессии могут быть связаны с аномальными изменениями активности главного циркадианного водителя ритма в СХЯ. Клеточные и молекулярные механизмы циркадианных ритмов в нейронах СХЯ в большей степени идентифицированы и регулируются генетически. Гены, кодирующие ключевые элементы циркадианных часов у млекопитающих, включают period (perl, реr2, реrЗ), neuronal PAS domain protein-2 (NPAS2), circadian locomotor output cycles kaput (СLOСК), cryptochrome (Cryl, Cry2) brain and muscle ARNT-like-1 (BMAL1) гены. Белки, кодируемые этими генами, являются частью циркадианной саморегулирующейся нейрональной петли, включая белки активаторы и супрессоры, активность которых также подчиняется суточному ритму и фактически задает ритм нейронам СХА (Laposky A. D. et al., 2006). У мышей носителей мутации CLOCK гена наблюдается поведение, напоминающее маниакальное состояние, включая гиперактивность, сокращение времени сна, редукцию страха, повышение подкрепляющей значимости кокаина, сукрозы и стимуляции медиального узла переднего мозга (Roybal К. et al., 2008). Интересно, что многие формы подобного маниакального поведения редуцируются после введения лития и возобновляются при локальной экспрессии функционального CLOCK протеина в вентральной теменной области СLOСК мутантной мыши (Roybal К. et al., 2008). В одном из исследований показано, что единичный нуклеотидный полиморфизм в PER2, NPAS2, mdBMALl генах связан с повышенным риском развития сезонной депрессии. Кроме того, определенные аллельные комбинации нуклеотидов в этих генах оказывают дополнительный эффект, повышая риск развития сезонного аффективного расстройства в 4,43 раза в сравнении с другими генотипами и 10,67 раз в сравнении с наиболее протективным генотипом (Partonen Т. et al., 2005).
Первичные или вторичные нарушения биологических часов на молекулярном уровне могут быть причиной нарушений эндогенных циркадианных ритмов, которые, в свою очередь, могут способствовать развитию депрессии. Другая точка зрения предполагает, что вместо или наряду с молекулярными аномалиями водителя ритма, изменения внешних факторов, задающих ритм (zeitgebers), могут вызывать депрессивные симптомы у биологически предрасположенных субъектов (Ehlers С. L. et al., 1988). Данная теория предполагает, что депрессивные эпизоды развиваются вследствие определенных событий жизни, вызывающих нарушения в социальных стимулах ритма, таких как время питания, распорядок работы, социальные потребности, интерперсональные отношения, которые в свою очередь, приводят к отклонениям в индивидуальных социальных ритмах. Это может нарушать способность организма сохранять стабильные биологические ритмы, особенно ритм
сон-бодрствование и ритмы психомоторной активности, бодрствования и аппетита. У большинства индивидов эти ритмы быстро стабилизируются после разрешения стрессовых событий, однако у предрасположенных субъектов они могут провоцировать развитие депрессии.
При депрессиях генетическая предиспозиция и разнообразные преморбидные факторы формируют нейробиологический субстрат, который фенотипически проявляется в неустойчивости биоритмической структуры организма и отражает снижение его компенсаторных возможностей по ресинхронизации системы. Другими словами, биоритмическая среда недостаточно инертна, чтобы быстро гасить естественно увеличивающуюся в ответ на какие-либо внешние воздействия амплитуду колебаний того или иного ритма.
В соответствии с получившей наибольшее распространение в мире схемой A. A. Borbely (1982) сон регулируется двумя находящимися в реципрокных отношениях процессами. Первый из них, процесс Ц, контролируется сильным Х-осциллятором, ритмическая активность которого мало зависит от внешних влияний и интимно связана с
гипоталамо-гипофизарно-адреналовой осью. Второй, процесс С, регулируется гомеостатически, т. е. более подвержен влиянию факторов внешней среды, и, возможно, целиком регулируется одним гипоталамическим нейропептидом, при достижении критической концентрации которого в крови (примерно через 2 суток) неизбежно наступает сон (Ursin R., 1984; Kupfer D., 1989). Именно этот процесс, эволюционно несущий адаптивную функцию, играет важную роль в формировании резерва компенсаторных возможностей по стабилизации биоритмической организации, поскольку сдерживает активность процесса Ц и гасит чрезмерный размах его колебаний.
Недостаточность процесса С является предпосылкой для развития заболевания. В этих условиях повторные социально-психологические стрессовые воздействия по известной схеме Селье через гипоталамо-гипофизарно-адреналовую систему, в том числе усиление утренних выбросов глюкокортикоидов (Post R. N., Weiss S. R., 1989) и кортикотропный релизинг-фактор (Nemeroff С. В., 1989), активизируют процесс Ц, который не возвращается полностью в исходное положение, как в норме. Остаточное усиление амплитуды колебаний и небольшие смещения ритмов постепенно суммируются до тех пор, пока компенсаторный резерв процесса С не истощается, что выражается в их десинхронизации, резком смещении циркадианных ритмов на более раннее время суток (опережение фазы).
Экспрессия большинства ритмов на поведенческом, физиологическом и биохимическом уровнях регулируется совокупностью сигналов из циркадианного осциллятора (процесса Ц) и информации о состоянии сон-бодрствование (процесса С) (Turek F.W., 2007). Поэтому центры контроля циркадианных ритмов и сна совместно участвуют в процессе временной координации внутренней и внешней среды для оптимизации выживаемости живых организмов. По-видимому, первичные циркадианные нарушения цикла сон-бодрствование приводят к нарушению сна, что вызывает десинхронизацию многих эндогенных ритмов и ведет или способствует развитию депрессии. Эта гипотеза подтверждается тем, что инсомния является фактором риска манифестации депрессивного расстройства, а также обострения и развития повторных эпизодов депрессии (Ford D.J., Kameron D.B., 1989; Chang Р. Р. et al., 1997; Dombrovski A. Y. et al., 2007). Большинство циркадианных аномалий, наблюдаемых при депрессии, корректируются после наступления ремиссии. Следовательно, нельзя исключить вероятность того, что эти аномалии являются только следствием депрессии. Однако, даже в этом случае десинхроноз симптомов может влиять на течение и прогноз заболевания.
Хронобиологические механизмы, в частности десинхронизация циркадианного ритма с циклом сон-бодрствование, играют важную роль в патогенезе депрессии, могут приводить к ее затягиванию, ускорению фазообразования и ухудшать прогноз заболевания в целом. Поэтому коррекция десинхроноза является первостепенной задачей в комплексной терапии РД.

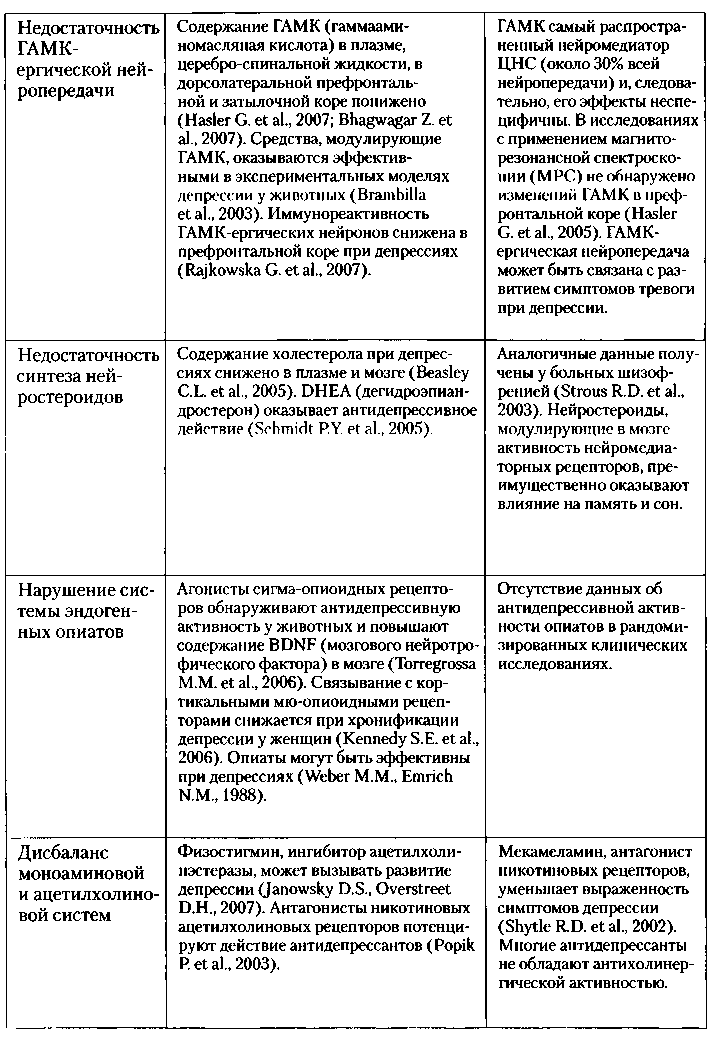
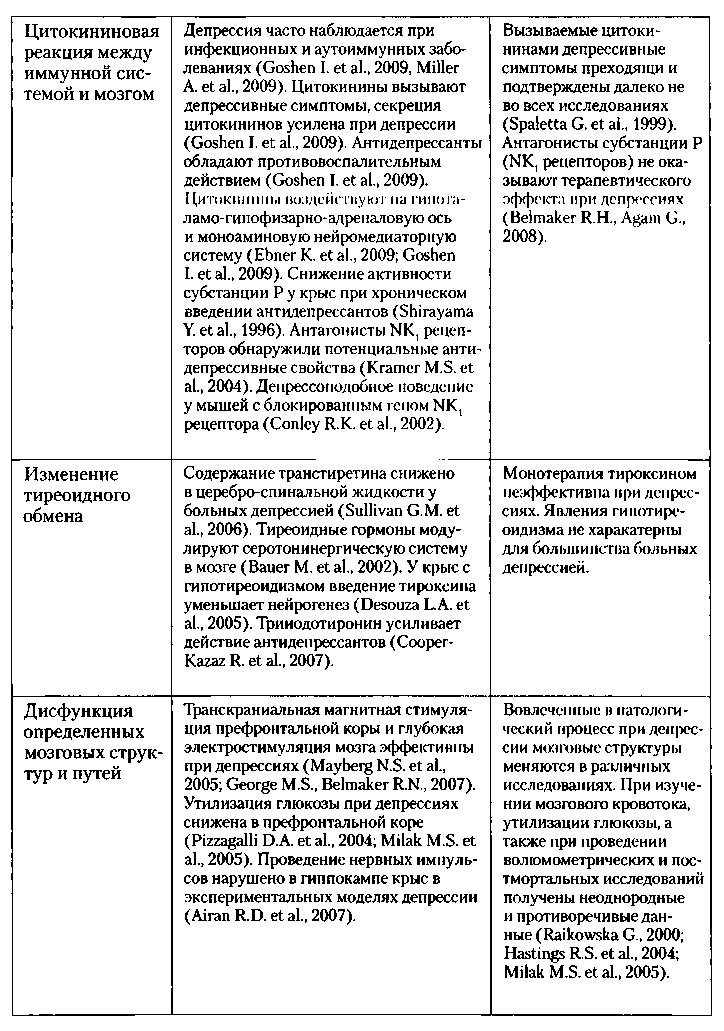
Таблица № 1. Другие биологические гипотезы патофизиологии депрессии