Основные сведения, классификация и краткие эпидемиологические данные
Клиническая онкология включает множество заболеваний, существенно различающихся своим течением, прогнозом и методами лечения в зависимости от происхождения опухоли, ее гистологической формы, локализации и других факторов.
Прогресс, достигнутый в последние годы в биологии, обусловлен теми чрезвычайными усилиями, которые предпринимаются мировым научным сообществом в борьбе со злокачественными новообразованиями. Причины канцерогенеза коренятся глубоко в самой основе живого и тесно переплетены с такими фундаментальными понятиями, как деление клетки, межклеточные взаимодействия, смерть, старение и бессмертие.
Только в последней четверти XX в. появились реальные предпосылки решения проблемы злокачественных новообразований. Главные из них — расшифровка структуры, функции и регуляции гена, дальнейшее развитие генной и клеточной инженерии и особенно компьютеризация научных исследований.
Достигнутый благодаря этому прогресс сопоставим по своему масштабу и значению с научными революциями начала XX в. в физике. Научная революция в биологии, свершаемая на наших глазах, еще далеко не завершена. В начале нынешнего века планируется окончание проекта «Геном человека», который направлен на расшифровку всех нуклеотидных последовательностей ДНК человека для проникновения в суть генетических программ, управляющих жизнью клетки и организма.
Интенсивность исследовании в этой области и их специализация чрезвычайно велики. В данной главе приведены лишь наиболее важные сведения об опухолевом росте.
Опухоль (новообразование, бластома. неоплазма) — патологическое разрастание, отличающееся от других патологических разрастаний (гиперплазия, гипертрофия, регенерация после повреждения) наследственно закрепленной способностью к неограниченному, неконтролируемому росту.
Существует два основных типа опухолей — доброкачественные и злокачественные.
Доброкачественные опухоли.
Такие опухоли растут, раздвигая прилежащие ткани, иногда сдавливая их, но обычно не повреждая; в некоторых случаях они инкапсулируются. Доброкачественные опухоли, как правило, не оказывают неблагоприятного воздействия на организм, поэтому их можно рассматривать как местные разрастания, не препятствующие отправлению жизненно важных функций. Их клиническое значение невелико. Исключение составляют лишь те случаи, когда сама локализация опухоли является фактором, угрожающим жизнедеятельности организма, например, при ее возникновении в головном мозге и сдавлении вследствие этого нервных центров.
Злокачественные опухоли.
Это многочисленная группа тяжелых, хронических заболеваний, заканчивающихся, как правило, летальным исходом, если отсутствовала или запоздала лечебная помощь. Злокачественные опухоли характеризуются инвазивным ростом, они инфильтрируют прилегающие ткани, образуют перифокальные очаги воспаления, часто метастазируют в близлежащие лимфатические узлы и отдаленные ткани, оказывают генерализованное воздействие на весь организм, расстраивая его гомеостаз. Все последующее изложение посвящено описанию именно этого типа опухолей.
Гистологические типы опухолей.
Организм человека состоит из клеток примерно 100 разных типов и почти все они могут трансформироваться в опухолевые. В зависимости от типа трансформированных клеток опухоли подразделяются на рак (происходят из клеток эпителия) и саркомы (происходят из клеток соединительной ткани). Поскольку первые возникают примерно в 10 раз чаше, чем вторые, термином «рак» зачастую пользуются для обозначения всех злокачественных новообразований. Вместе с тем благодаря повсеместному присутствию в организме соединительнотканных элементов саркомы могут появиться практически в любом органе или ткани.
Локализация и гистологический тип опухоли во многом определяют скорость ее роста, чувствительность к тем пли иным терапевтическим воздействиям, способность давать метастазы и рецидивы и в конечном итоге клиническое течение и прогноз. Поэтому гистологический диагноз опухоли имеет первостепенное значение для выбора стратегии лечения.
Существует много форм рака, например, аденокарцинома (рак железистый — возникает из эпителия желез), рак папиллярный (образует сосочковые структуры), бронхиоло-альвеолярный (из эпителия бронхов), рак плоско клеточный, перстневидноклеточный, овсяноклеточный, мелкоклеточный, гигантоклеточный (по форме образующих их клеток), рак медуллярный (по внешнему сходству с тканью мозга), скирр («твердый» рак с преобладанием стромальных элементов), рак эпидермоидный (по сходству с многослойным плоским эпителием кожи) и т.д.
Саркомы подразделяются (по своей локализации) на саркомы костей, мягких тканей и органов, а по типу исходных клеток — на фибросаркомы, липосаркомы, лейомиосаркомы и рабдосаркомы (происходят из мышечных элементов), а также лимфосаркомы, хондросаркомы и т.д.
Распространенность.
Опухолевые заболевания, поражающие всех представителей животного мира, настолько широко распространены, что они являются второй после сердечно-сосудистых заболеваний причиной смертности. В современном мире примерно каждый четвертый человек сталкивается с онкологической патологией, и каждый пятый погибает по этой причине. Для младенца, родившегося в России в 1992 г., вероятность заболеть злокачественным новообразованием на протяжении предстоящей жизни составляет 19,6% для мальчика и 16,0 % для девочки, а вероятность умереть от этой патологии — 16,5% для мальчика и 10,8 % для девочки. Число онкологических заболевании растет, что объясняется как общим старением населения, так и увеличением влияния канцерогенных факторов. Все чаще злокачественные опухоли возникают в детском и младенческом возрасте.
Факторы риска, способствующие развитию опухоли.
Эти факторы, обычно называемые факторами риска, подразделяют на три основные группы: вредные привычки, плохие условия труда, загрязнение окружающей среды.
Наиболее распространенная вредная привычка — табакокурение, которое является причиной 90 % случаев рака легкого, особенно у мужчин, и менее часто — причиной других форм опухолей: желудка, полости рта, глотки и печени.
Риск заболевания злокачественными новообразованиями увеличивается у людей, находящихся на диете, богатой животными жирами и копчеными продуктами, с малым содержанием клетчатки. Очень важную роль играют высокая концентрация нитратов и пестицидов в пище и воде, резко повышающих риск развития опухолей. И наоборот, витамины С, А, β-каротин, особенно в овощах и фруктах, оказывают защитный эффект. Чрезмерный загар повышает риск возникновения меланом. До 4 °с злокачественных новообразований связано с профессиональной деятельностью. В 1897 г. был впервые описан рак мошонки у трубочистов в Лондоне.
Перечень вредных производств все увеличивается, включая производство анилиновых красителей, асбеста, асфальта, инсектицидов, фармакологических препаратов и др. Более 100 веществ, производимых человеком, являются канцерогенными. Эти вещества вместе с промышленными отходами загрязняют водоемы, атмосферу: они находятся в составе строительных материалов, попадают в пищу. Немаловажную роль играют и радиоактивные соединения. В последние годы интенсивно изучают роль различных факторов в развитии злокачественных новообразований. По данным научной литературы, 30 % всех случаев заболевании раком связаны с табакокурением, 3% — с употреблением алкоголя, 35 % с неправильным питанием, 5 % ~ с профессиональными вредностями. в том числе с отходами промышленного производства.
Генетическая предрасположенность.
Явная генетическая предрасположенность к опухолям в так называемых «раковых семьях» встречается в 5—10 % случаев злокачественных новообразований. Наиболее высокая частота возникновения злокачественных опухолей (до 100%) и относительно ранний возраст заболевших, связанных родственными отношениями, бывают при наследственных заболеваниях. К ним относятся: синдромы Ли —Фраумени, Гарднера и Блума, пигментная ксеродерма, анемия Фанкони, семейный полипоз кишечника, атаксия-гелеангиэктазия и многие другие.
Существенный прогресс в понимании роли наследственности в происхождении злокачественной опухоли связан с завершением проекта «Геном человека». Расшифрована почти полностью последовательность трех миллиардов нуклеотидов, составляющих хромосомную ДНК человеческой клетки. Это — одно из значительных достижений минувшего столетия.
Вместе с тем научный прогресс, свершаемый на наших глазах, еще далеко не завершен. Немало, в частности, времени потребуется для того, чтобы извлечь из расшифрованных генетических текстов их функциональное содержание. От этого в определенной степени зависит глубина нашего познания нормального устройства клетки и механизмов ее злокачественной трансформации.
В структуре общей заболеваемости рак в челюстно-лицевой области составляет 32,5% (Н. Н.Трапезников и соавт., 1997; А. И.Пачес, 1997). Рак нижней губы составляет 3 — 8%, рак языка — около 55%. щеки — 12 — 15%, дна полости рта — 10—12%, альвеолярных отростков верхней челюсти и твердого нёба — 5—6 %, альвеолярного отростка нижней челюсти — 5 — 6 %, мягкого нёба — 6 — 7 % (П. Г. Битюцкий и соавт., 1996).
Рак органов полости рта среди опухолей головы и шеи занимает второе место после рака гортани и в 5 —7 раз чаще развивается у мужчин, чем у женщин. Предрасполагающими факторами возникновения рака губы и органов полости рта являются неблагоприятные экологические и атмосферные воздействия и, по-видимому, отягощенная наследственность. Хронические механические травмы и раздражения, табакокурение, атрофия покровного эпителия в пожилом возрасте, злоупотребление алкоголем также способствуют появлению предраковых заболеваний.
Наиболее частым предраковым заболеванием красной каймы губы является хейлит Манганотти (34,9 %) и ограниченный гиперкератоз (25,5 %). Что касается предракового поражения слизистой органов полости рта, то слизистая языка поражается чаще лейкоплакией в различных вариантах (38,3 %). То же относится и к слизистой оболочке дна полости рта, но частота поражения при этом отмечена значительно больше в сравнении с поражением других органов и составила (57 %).
Отмечено, что рак губы и слизистой оболочки полости рта встречается значительно чаще у мужчин (81,1 %), чем у женщин (18,9 %). Возраст больных составляет в среднем от 51 до 70 лет с тенденцией к возникновению поражения в более молодом возрасте.
Этиологические факторы канцерогенеза
В подавляющем большинстве случаев опухоли у человека появляются «спонтанно», т.е. без видимых причин. Иногда на фоне полного здоровья этот диагноз ставят пациенту в результате случайного обследования. Чаще опухоль дает о себе знать медленно, но неуклонно нарастающей симптоматикой — различной для опухолей разного происхождения и локализации. Невозможно выявить в каждом отдельном случае, что явилось этиологической причиной заболевания.
По мере выяснения механизмов канцерогенеза становится все более очевидным, что, во-первых, возникновение опухоли — процесс многостадийный (причем каждый последующий «шаг» может быть спровоцирован разными факторами) и, во-вторых, что почти все канцерогены генотоксичны, т.е. вызывают повреждения ДНК (давно бытует убеждение, что «рак — это болезнь генов»). Канцерогены повреждают ДНК случайно (т.е. неспецифичны в отношении ее нуклеотидных последовательностей), но при возрастании дозы канцерогенного воздействия растет, естественно, и вероятность того, что в одной из десятков триллионов клеток организма окажутся задетыми гены, управляющие делением клетки. Поскольку и в окружающей, и во внутренней среде организма существует множество канцерогенов, то в большинстве случаев нельзя выявить конкретный этиологический фактор начала опухолевого роста.
Мнение о канцерогенных факторах как генотоксических агентах является наиболее распространенным и благодаря этому представление о химическом канцерогенезе многие предпочитают вирусной и физической теории возникновения злокачественного роста. Теперь ясно, что этиологических факторов много, тогда как патогенез (в принципиальном плане) — един и сводится к нарушению механизма регуляции клеточного деления на одном из его многочисленных этапов.
Вирусный канцерогенез.
Вирусы, вызывающие опухоли у животных, подразделяют на ДНК-содержашие (например, вирус обезьяны SV40) и РНК-содержащие, или ретровирусы (например, вирус саркомы Рауса).
Этиологическая роль вирусов в возникновении опухолей человека весьма вероятна в случае лимфомы Беркитта, рака носоглотки (ДНК-содер-жаший вирус Эпштейна—Барр), рака шейки матки (вирус папилломы), а также Т-клеточного лейкоза взрослых (ретровирус HTLV-1). В отношении некоторых других опухолей человека роль вирусов только предполагается.
Химический канцерогенез.
Впервые японские ученые Ямагива и Ишикава вызвали рак у кроликов с помощью каменноугольной смолы в 1915 г. В настоящее время известно около 20 химических канцерогенов, способных вызвать опухоли у человека, — промышленные. лекарственные, естественного происхождения.
Широко распространена в окружающей среде многочисленная группа полициклических ароматических углеводородов (ПАУ). Основным ее представителем является бенз(а)пирен, который образуется не только в результате производственной деятельности человека, но и представляет собой природный канцероген — он содержится в почве, выбросах вулканов. К ПАУ относятся метилхолантрен, диметилбенз(а)антрацен и др.
В 100 сигаретах содержится 1,1 — 1,6 мкг бенз(а)тирена, а примесь мышьяка в 15 раз превышает его максимально допустимое количество. В газовой фазе и твердых частицах табачного дыма находятся дибенз(а)антрапен и никель, канцерогенный для человека. У людей, выкуривающих 16— 25 сигарет в день, риск заболеть раком в 30 раз выше, чем у некурящих. Считается, что у мужчин США в 85 — 90 % случаев заболевание раком легкого вызвано курением сигарет.
Канцерогенные нитрозосоединения вызывают опухоли у разных видов животных. Нитрозометил и нитрозоэтил мочевины, нитрозодиметиламины могут синтезироваться в организме из нитритов (нитратов) и вторичных аминов, содержащихся в пище. В толстой кишке при участии бактериальной флоры образуются вторичные амины, и, кроме того, нитраты переводятся в нитриты.
Большинство химических канцерогенов относят к проканцерогенам. Они превращаются в истинные, конечные канцерогены только после метаболических превращений, катализируемых тканевыми ферментами (неспецифическими окемдазами), локализованными в эндоплазматическом ретикулуме и частично в ядре клетки. Полициклические ароматические углеводороды типа бенз(а)пирена или диметил -бенз(а)антрацена становятся конечными канцерогенами, превращаясь в эпоксиды. Некоторые про канцерогены становятся конечными в результате спонтанных реакций.
Меньшую группу составляют прямые канцерогены, например, нитрозамины, р-пропионлактон, диметил-карбамилхлорид; они действуют как таковые, не подвергаясь метаболической модификации.
Все химические канцерогены реагируют с клеточной ДНК, ковалентно присоединяясь к ней, они образуют многочисленные аддукты, индуцируют одно- и двунитевые разрывы.
Трансплацентарный канцерогенез.
Механизм этого концерогенеза состоит в том, что у детенышей экспериментальных животных спустя несколько месяцев после рождения появляются опухоли в результате действия в период их внутриутробного развития малых (безвредных для матери) доз химических канцерогенов. Эффект достигается и в том случае, когда канцероген попадает в эмбрион непосредственно, не будучи модифицированным тканевыми ферментами матери. У таких детенышей возникают опухоли различных локализаций, в том числе в головном мозге. Нервная система зародыша крысы в десятки раз чувствительнее к канцерогену, чем у взрослого животного.
Человек также должен считаться с этим явлением. Обнаружены случаи возникновения светлоклеточного рака влагалища у девочек и девушек, чьи матери во время беременности принимали диэтилстилбэстрол. В странах Западной Европы повышается частота онкологических заболеваний подростков (лейкозы, опухоли нервной системы, почек). Умножаются случаи смерти от этих видов рака среди детей моложе 15 лет.
Радиационный канцерогенез.
Канцерогенное действие ионизирующего излучения обнаружено в 1902 г., т.е. вскоре после открытия рентгеновских лучен и естественной радиоактивности. Радиацией можно индуцировать опухоли практически всех органов, чаше всего кожи и костей, лейкозы, а также железистой ткани.
Частота и виды злокачественных новообразований, индуцированных ионизирующими излучениями, зависят от проникающей способности излучения, характера воздействия (внешнее или внутреннее), органотропности радионуклидов и т.д.
Ультрафиолетовое излучение.
Длительное воздействие ультрафиолетового излучения вызывает меланомы на открытых участках кожи (голова, шея, руки). В этом отношении наиболее чувствительны блондины со светлой кожей.
Характерные свойства опухолей
Злокачественные опухоли характеризуются катаплазией, метаплазией, дисплазией, инвазивным и деструктивным ростом метастаз.
Катаплазия (анаплазия). Термин обозначает снижение уровня дифференцировки, появление слабодифференцированных или недифференцированных клеток, похожих на эмбриональные. Опухоль может утрачивать частично или полностью тканеспецнфические признаки. Этот процесс протекает хаотично и нередко приводит к образованию атипичных клеток, не имеющих аналогов в нормальных тканях.
Метаплазия (от лат. metaplasis — преображение). Это стойкое изменение морфофизиологических свойств клеток (ткани), сопровождающееся превращением их в клетки (ткань) другого типа (например, клетки соединительной ткани начинают образовывать кость в неподходящем месте).
Дисплазия (от лат. dys — нарушение, расстройство и plasis — форма, образование). Данный термин обозначает нарушение в опухолевом очаге характерной для данной ткани структуры, ее атипию (по строению, расположению и взаимоотношениям клеточных элементов опухоль резко отличается от исходной нормальной ткани).
Инвазивный и деструктивный рост, метастазирование.
Опухоли растут, инфильтрируя (прорастая) окружающие ткани и вызывая их деструкцию. Они часто дают метастазы (вторичные очаги в отдаленных тканях и органах), что во многих случаях означает финальную стадию процесса.
Входе метасгазирования отбирается и выживает небольшая субпопуляция клеток, предсуществующих в «родительской» опухоли. Способность к метастазированию зависит как от свойств самих опухолевых клеток, так и от условий внутренней среды организма. Отбор опухолевых клеток для метастазирования является многоэтапным процессом, который схематически можно представить следующим образом:
первичный очаг → новообразование сосудов → прорастание в сосуды (лимфатические, капиллярные, венозные) → образование эмболов (многоклеточных агрегатов, включающих лимфоциты и тромбоциты) → транспорт в отдаленные ткани и органы → задержка в капиллярах → прикрепление → экстравазация (выход из сосудов) → адаптация к микроокружению → пролиферация → формирование метастаза.
В росте опухоли и ее метастазов важную роль играет ангиогенез. От холидиаметром 1—2 мм могут питаться посредством диффузии, а увеличение их размеров возможно только при адекватном кровоснабжении. Благодаря продукции факторов ангиогенеза в опухолевый очаг мигрируют эндотелиальные клетки из прилегающей соединительной ткани. Эти клетки размножаются, что обусловливает врастание сосудов. Активность ангиогенеза в опухоли определяется соотношением активаторов (ангиогенин. фактор роста тромбоцитов, трансформирующие ростовые факторы а и β, фактор некроза опухолей, простагландины Е1 и Е2. интерлейкин-8 и др.) и ингибиторов (ангиостатин, ингибитор хрящевой ткани, гепариназа, интерфероны α и β, тромбоспондин, тканевый ингибитор металлопротеиназ и др.).
Преобладание ингибиторов ангиогенеза нарушает питание опухоли и приостанавливает ее рост, переводя в «дремлющее» состояние. В центре опухоли при этом часто обнаруживают участки некроза. И, наоборот, при улучшении кровоснабжения оставшиеся в живых раковые клетки могут дать начало рецидиву опухоли даже спустя несколько лет после удаления основного очага.
В крови онкологических больных, не имеющих метастазов, часто обнаруживают циркулирующие опухолевые клетки, которые разрушаются естественными киллерами и макрофагами. Выживает менее 0,05 % клеток благодаря их естественному отбору на устойчивость к природным «киллерам». Сохранившие жизнеспособность клетки задерживаются в узких сосудах того органа, к которому они имеют тропность (многие опухоли проявляют тенденцию к преимущественному метастазированию в определенные ткани, например., аденокарцинома молочной железы метастазирует в кости и головной мозг, а нейробластома — в печень и надпочечники).
Взаимоотношения опухоли и организма
Организм является для опухоли внешней средой, с одной стороны, создающей необходимые условия для ее существования и роста, а с другой противодействующей ее развитию. Развитие опухоли — интерактивный процесс (акты «агрессии» со стороны опухоли чередуются с ответными «контрмерами» организма). Исход этой борьбы предопределен громадным потенциалом «агрессивности» опухоли, с одной стороны, и ограниченностью защитных ресурсов организма — с другой.
Иммунная защита.
Образовавшийся в организме клон трансформированных клеток дает начало злокачественной опухоли только при определенных условиях. Первым барьером для роста опухоли является система «естественной неспецифической резистентности», способная элиминировать небольшое число опухолевых клеток. Она представлена естественными киллерами — крупными гранулярными тимуснезависимыми лимфоцитами, число которых составляет от 1 до 2,5% всей генерации периферических лимфоцитов. Они активируются интерфероном, могут лизировать не только опухолевые, но и нормальные поврежденные клетки. Определенной противоопухолевой активностью обладают макрофаги, благодаря генерации кислородных радикалов и пероксида водорода. Устойчивость опухолевых клеток к факторам естественной резистентности зависит от их способности продуцировать факторы, подавляющие эту систему (простагландины Е), и их устойчивости к Н2О2.
Специфический противоопухолевый иммунитет формируется на поздних стадиях опухолевого роста и мало активен. Это обусловлено, как правило, слабой антигенностью опухолей. Редкие случаи регрессии злокачественных опухолей (меланомы, нейробластомы у детей) указывают, однако, на принципиальную возможность успешной борьбы организма с уже сформировавшейся неоплазмой.
Системное действие опухоли на организм.
Прогрессирующий опухолевый процесс в конечном счете приводит организм к гибели вследствие различных причин. Часто возникают тромбоэмболии или кровотечения, поражения жизненно важного органа (головного мозга), резко нарушается гормональный баланс при опухолях эндокринных желез, секретирующих биологически активные вещества.
Как правило, на начальных этапах развития опухоль слабо себя проявляет, что затрудняет ее раннюю диагностику.
Опухоль как «ловушка» питательных веществ.
Интенсивный рост опухоли требует ее пластического и энергетического обеспечения, что достигается особенностями изоферментного спектра, который обеспечивает накопление и усвоение опухолевыми клетками различных питательных веществ, вызывая в организме состояние их дефицита (аминокислот, глюкозы, витаминов и др.). Опухоль извлекает азотистые вещества не только из пищи, но и из продуктов распада тканевых белков: мышечной, лимфатической и других, что характеризует паразитирование опухоли, т.е. увеличение ее массы за счет уменьшения массы тела. В целом этот механизм отражает общее катаболическое действие веществ, продуцируемых опухолевой тканью.
Опухоль называют «ловушкой» глюкозы, которую она использует для синтеза белков и нуклеиновых кислот. В результате создается дефицит глюкозы в организме. Гипогликемия вызывает включение механизмов компенсации — гликонеогенез, т.е. образование глюкозы из белков и жиров. Гликонеогенез инициируется глюкокортикоидами, которые восполняют запасы глюкозы за счет распада тканевых белков, в том числе лимфоидной ткани, что вместе с другими факторами ослабляет иммунитет. К тому же опухоль усваивает большое количество витаминов и других жизненно важных веществ.
Опухоль как источник биологически активных соединений.
Опухолевая ткань, создавая, с одной стороны, дефицит определенных соединений, способна, с другой стороны, продуцировать разнообразные биологически активные вещества, не свойственные нормальному организму, изменяющие обмен веществ и оказывающие токсический эффект. К ним относятся продукты распада опухоли, которые образуются, прежде всего, вследствие некроза центральной ее части из-за недостаточной васкуляризации. Продукты распада вызывают системную неспецифическую реакцию, заключающуюся в обшей интоксикации, лихорадке, одышке и др.
Специфическое воздействие секретируемых опухолью веществ связано с различными ростовыми и ангиогенными факторами. Они способствуют размножению клеток отдаленных от опухоли тканей, если на клетках есть соответствующие рецепторы. Некоторые злокачественные опухоли секретируют также несвойственные данной ткани «эктопические» гормоны, вызывающие общий дистантный эффект. Например, ткань рака легкого нередко секретирует адренокортикотропный гормон или инсулин, глюкагон, а ткань рака молочной железы — гормон, имеющий сходство с секретом околощитовидной железы. Наибольшее воздействие на организм оказывают гормоны, секретируемые опухолями эндокринных желез.
Раковая кахексия (общее истощение организма).
Такое состояние развивается обычно в терминальном периоде опухолевой болезни. Кахексия обусловлена усиленным распадом белков скелетных мышц, частично миокарда, а также истощением жировых депо. Сопровождается отвращением к пище (анорексией) и изменением вкусовых ощущений. Причиной кахексии являются повышенный (иногда на 20 — 50 %) расход энергии вследствие гормонального дисбаланса (преобладание катаболических гормонов) и общая интоксикация.
В сыворотке крови больных, страдающих хроническими заболеваниями и злокачественными новообразованиями. нередко обнаруживают кахектин — цитотоксический полипептидный гормон, известный также как TNF (tumor necrosis factor, фактор некроза опухолей), секретируемый макрофагами. Системное действие TNF связано с тем, что все клетки организма обладают рецепторами к этому фактору, при высокой концентрации которого развиваются различные эффекты: шоковое состояние, падение кровяного давления, расстройства липидного и углеводного обмена, метаболический ацидоз, анорексия, кахексия, активация нейтрофилов, вплоть до гибели организма. Фактор некроза опухоли вызывает аналогичные эффекты и при других неопухолевых хронических заболеваниях, сопровождающихся интоксикацией.
Биохимические показатели.
Опухоль вызывает значительные нарушения обмена веществ в организме больного, что проявляется в изменении соответствующих биохимических показателей. Это имеет диагностическое и прогностическое значение и позволяет в ряде случаев судить о локализации, степени распространения опухоли, функциональном состоянии жизненно важных органов.
Для оценки синтетической функции печени определяют: общий белок, альбумин, билирубин, активность аланиновой трансаминазы; функции почек — азотистые вещества; состояние костной ткани и минерального обмена.
Опухолевые маркеры.
Это соединения, обнаруживаемые в биологических жидкостях онкологических больных и синтезируемые либо собственно раковыми клетками, либо клетками нормальных тканей в ответ на инвазию опухоли. Маркерами опухоли могут быть ферменты, гормоны, антигены (внутриклеточные или ассоциированные с поверхностными мембранами) и метаболиты. Концентрация маркеров в среде часто коррелирует с массой опухоли и ее пролиферативной активностью.
Появление маркеров обусловлено особенностями метаболизма раковой клетки. Так, опухоль может утратить некоторые изоферменты, присутствующие в гомологичных нормальных тканях, и, напротив, продуцировать изоформы, характерные для данной ткани только в период ее эмбрионального развития. В опухоли могут меняться активность лизосомальных и мембраносвязанных ферментов, синтезироваться эктопические изоэнзимы и гормоны. Необходимо, однако, подчеркнуть, что ни в трансформированных клетках, ни в биологических жидкостях онкологических больных не обнаружены такие соединения, которые были бы характерны только для опухоли и не обнаруживались бы в нормальных тканях на тех или иных стадиях их развития.
Современные методы позволяют выявить опухолевые маркеры в столь малых (фетамолярных, М) концентрациях, что это дает возможность в ряде случаев следить за ходом заболевания и эффективностью лечения. Опухолевые маркеры можно подразделить на две основные группы: продуцируемые самой опухолью и ассоциированные с опухолью (их появление в последнем случае обусловлено опухолевым процессом независимо от того, какой тканью они продуцируются).
Среди маркеров, продуцируемых опухолью — альфа-фетопротеин, его содержание в сыворотке крови повышается при гепатоцеллюлярном раке; раково-эмбриональный антиген у больных раком толстой кишки, поджелудочной железы, молочной железы и легкого; тканевый полипептидный антиген при раке мочевого пузыря, предстательной железы и почек; хорионический гонадотропин (при опухолях трофобласта).
Маркеры, ассоциированные с опухолью, — это белки острой фазы воспаления (церулоплазмин, гаптоглобин, α2-глобулины, С-реактивный белок), некоторые ферменты, иммунные комплексы. Содержание этих белков неспецифически повышается в ответ на другие патологические процессы (воспаление, травмы и др.).
При различных онкологических заболеваниях отмечено повышение активности сывороточных ферментов — лактатдегидрогеназы (ЛДГ) и креатинкиназы, которые поддерживают оптимальные концентрации АТФ и АДФ в клетке. Повышение активности креатинкиназы отмечается чаще всего у больных с опухолями желудочно-кишечного тракта.
Стадии развития опухоли
Выделяют три основные стадии канцерогенеза — инициация, промоция и прогрессия.
Инициация.
Это начальный этап в длинной цепи событий, ведущих к образованию опухолевого очага. Инициация заключается в мутации одного из генов, регулирующих клеточное размножение. Клетка становится инициированной, т.е. потенциально способной к неограниченному делению, но требующей для проявления этой способности дополнительных условий. Инициирующими факторами являются различные канцерогены, вызывающие повреждение ДНК.
Промоция.
К промоторам относятся химические вещества, которые не вызывают повреждения ДНК, т.е. не являются канцерогенами, но при длительном воздействии на инициированные клетки способствуют возникновению опухоли. По-видимому, промоторы стимулируют клеточное деление, что создает критическую массу инициированных клеток и способствует их высвобождению из-под тканевого контроля.
Прогрессия.
Рост опухоли — это не только увеличение числа однородных клеток. Опухоль постоянно претерпевает качественные изменения и приобретает новые свойства — все большую автономность от регулирующих воздействий организма, деструктивный рост, инвазивность, способность к образованию метастазов (обычно отсутствующую на ранних этапах) и, наконец, ее поразительную приспособляемость к меняющимся условиям.
Признаки злокачественности возникают в ходе прогрессии независимо друг от друга, что объясняет разнообразие опухолевых фенотипов в отличие от нормально развивающейся ткани, структура и функция которой всегда четко определены.
Клональное происхождение опухолей.
Опухоль является потомством, т.е. генерацией одной первичной клетки, которая в результате многостадийно го процесса приобрела способность нерегулируемого роста.
Распространение опухоли по всему организму (метастазирование) также является следствием размножения генерации (клона) первично трансформированной клетки.
Следует отличать от метастазов первично множественные опухоли (несколько независимо возникших опухолей у одного больного). В этих случаях чаще всего есть генетическая предрасположенность к злокачественным новообразованиям.
Клональная гетерогенность опухолей.
Геном опухолевой клетки нестабилен, поэтому в процессе многократного деления появляются новые клоны, различающиеся генотипически и фенотипически. Это свойство определяет прогрессию опухоли, т.е. появление в результате естественного отбора клонов, обладающих быстрым ростом и устойчивостью к факторам защиты организма. Опухоль становится все более агрессивной, автономной и устойчивой к лечебным воздействиям.
Таким образом, непрерывно прогрессируя, претерпевая качественные изменения и проходя через необратимые стадии, опухоль «дрейфует» в направлении увеличения злокачественности.
Свойства опухолевых клеток
Свойства трансформированных меток можно изучать in vitro, что обусловлено их способностью легко переходить в культуру, сохраняя опухолевый фенотип. Благодаря широкому использованию различных клеточных линий оказалось возможным выяснить молекулярные механизмы канцерогенеза.
Изменения кариотипа и хромосомные аберрации.
Одно из наиболее характерных свойств трансформированных клеток — разнообразные изменения кариотипа (анэуплоидия, гиперплоидия), хромосомные аберрации (делеции, транслокации, амплификации) и мутации генов, а также присутствие экстрахромосомных элементов, вовсе не встречающихся в нормальных клетках. В опухолевом очаге, как правило, обнаруживаются клетки с существенно различающимся кариотипом, что отражает одно из самых характерных свойств опухолевой клетки — нестабильность ее генома.
При цитогенетическом анализе часто обнаруживается специфическая связь между изменением кариотипа и определенным типом опухоли: например, делеция длинного плеча 13-й хромосомы при ретинобластоме, появление так называемой «филадельфийской» хромосомы (перенос участка длинного плеча 9-й хромосомы на длинное плечо 22-й хромосомы) — при миелоидном лейкозе. Эти данные позволили предположить, что в этих участках (локусах) хромосом в норме имеются гены, тормозящие клеточное размножение, и их повреждение (или устранение) способствует развитию опухолевого процесса.
Признаки клеточной трансформации.
Трансформированные клетки, культивируемые in vitro, отличаются от нормальных клеток определенными свойствами.
- Они способны расти в культуре при низкой концентрации сыворотки и в отсутствие ростовых факторов; их размножение ограничено, по-видимому, лишь питательными ресурсами среды.
- Постоянно пребывают в цикле деления (не переходя в фазу покоя); остановка деления происходит только из-за отсутствия питательных веществ.
- Могут расти в суспензии, не испытывая потребности в прикреплении к внеклеточному матриксу.
- Способны неограниченно делиться вследствие отсутствия у них так называемого контактного торможения. что позволяет им образовывать при размножении многослойную культуру (нормальные клетки, соприкоснувшись своими поверхностями, прекращают делиться, в результате чего они образуют однослойную культуру).
- У трансформированных клеток ослаблены адгезивные свойства — они утрачивают способность распластываться на подложке и прочно прикрепляться к твердому субстрату; они приобретают способность размножаться в полужидкой среде.
- Главным отличительным признаком опухолевых клеток является их туморогенность — способность образовывать опухоли при имплантации экспериментальным животным.
- Иммортализация опухолевых клеток. Нормальные клетки в культуре in vitro не могут существовать длительно, поскольку после определенного числа клеточных делений скорость роста постепенно замедляется, затем прекращается полностью и клетки в конце концов погибают (этот феномен называют барьером Хейфлика). Число делений клеток в культуре различно у разных видов и зависит от возраста животного, от которого эти клетки были исходно получены (чем моложе организм, тем больше возможно число пассажей). Следовательно, нормальной клетке присуще заданное программой число делений, после чего наступает ее гибель апоптоз.
Трансформированные клетки в культуре могут делиться неограниченно долго, годами и десятилетиями, не проявляя признаков старения, т.е. у них имеются дефекты механизма апоптоза и поэтому они способны выживать в условиях, гибельных для нормальных клеток. - Важную сдерживающую роль по отношению к трансформированным клеткам могут играть их соседи. Роль межклеточной кооперации (и межклеточных контактов как механизма ее реализации) в функционировании нормальных клеток чрезвычайно велика. «Асоциальность» трансформированных клеток является, в частности, следствием ослабления их контактов с другими клетками. В клеточных ассоциациях существует баланс механизмов, стимулирующих и тормозящих деление, благодаря которому поддерживается тканевый гомеостаз. Эти межклеточные взаимодействия какое-то время сдерживают первично трансформированные клетки. Однако по достижении последними некой критической массы они способны «ускользнуть» из-под тканевого контроля и дать начало опухоли.
Опухолевые клетки приобретают черты «асоциальности» вследствие ослабления их контакта с другими, окружающими клетками, которые оказывали бы при нормальных межклеточных контактах действие, проявляющееся в нормализации опухолевого фенотипа.
Молекулярные механизмы клеточного деления
Опухолевый рост является следствием нарушения тканевого гомеостаза, поддерживаемого балансом клеточной пролиферации и гибели (апоптоза). Увеличение клеточной массы опухоли может быть обусловлено как усилением пролиферации, так и угнетением апоптоза. Вероятность «сбоев» в механизмах поддержания этого гомеостаза вполне реальна в условиях канцерогенного воздействия факторов внешней среды.
Наследуемые нарушения механизмов тканевого гомеостаза обусловлены теми или иными повреждениями структуры ДНК.
Нормальная делящаяся клетка с поврежденной ДНК либо прекращает деление до полной репарации повреждений, либо самоуничтожается (апоптоз). Последний вариант предпочтительней, поскольку потеря клетки может не иметь никаких последствий. Сохранение такой клетки таит для организма смертельную угрозу возникновения клона дефектных (потенциально опухолевых) клеток. Опухолевый рост возможен лишь тогда, когда дефектные клетки способны «проскальзывать» через защитный барьер апоптоза.
Ниже приведено краткое описание нарушений, обусловливающих трансформацию нормальных клеток в злокачественные.
В организме существуют два типа физиологической регуляции клеточного размножения — эндокринная и паракринная (рис. 12.1). Регуляторные молекулы секретируются клеткой и действуют вовне (через кровоток, на соседние клетки или себя). Утолщенные полукружия, изображенные на поверхности клеточной мембраны, — рецепторные участки.
Эндокринная регуляция.
Она осуществляется железами внутренней секреции (гипофиз, надпочечники, щитовидная, околощитовидная, поджелудочная и половые железы). Они секретируют продукты своей активности в кровь и оказывают генерализованное воздействие на весь организм.
Паракринная регуляция.
В отличие от эндокринной паракринная регуляция заключается в том, что секретаруемые клетками активные вещества распространяются за счет диффузии и действуют на соседние клетки-мишени. Так действуют, например, митогенные стимуляторы (полипептидные ростовые факторы) — эпидермальный фактор роста, фактор роста тромбоцитов, интерлейкин-2 (фактор роста Т-клеток), фактор роста нервов и т.д.
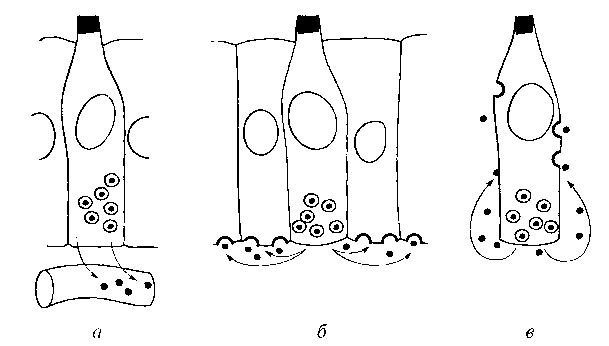{kind=link}
Аутокринная регуляция.
Она отличается от паракринной регуляции тем, что одна и та же клетка является источником ростового фактора и его мишенью. В результате может возникать феномен непрекращающегося, самоподдерживающегося митогенного «возбуждения» клетки, приводящего к нерегулируемому размножению. Клетка не нуждается во внешних митогенных стимулах и становится полностью автономной. С помощью аутокринной регуляции можно объяснить механизмы канцерогенеза. Для этого вначале разъясним понятие, называемое митогенной «рефлекторной дугой».
{kind=link}
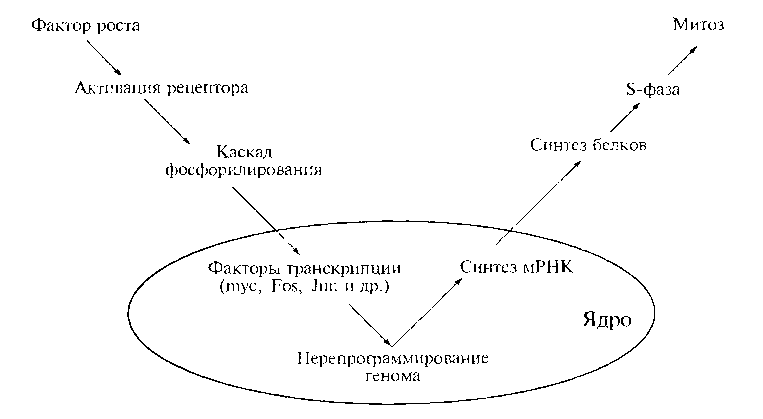
В регуляции сложных систем, как бы различны они не были, обнаруживаются общие черты. Между рефлекторной активностью организма и митотической активностью клетки есть принципиальное сходство (рис. 12.2).
Суть заключается в том, что на периферии системы (организм, клетка) существуют различные специализированные рецепторы (глаз, ух о, тактильные и обонятельные — в первом случае; рецепторы ростовых факторов — во втором); воспринимаемые ими внешние сигналы передаются внутрь системы (в виде импульсов по чувствительным нервам или в виде каскадов реакций фосфорилирования); затем происходит обработка сигнала в центре (центральной нервной системе пли в клеточном ядре) и информация центробежно (в виде импульсов по двигательным нервам или с помощью молекул мРНК) поступает к исполнительным органам и индуцирует их активность (двигательную, секреторную — в первом случае и митотическую — во втором).
Перенос митогенного сигнала от периферии клетки к ее ядру осуществляется в виде каскада реакций фосфорилирования посредством протеин-киназ (ферментов, фосфорилирующих белки). Существует три типа протеинкиназ (тирозиновые, сери новые и треониновые) по их способности фосфорилировать определенные аминокислоты. Фосфатные группы играют роль молекулярных переключателей: меняя конформацию определенных белковых структур (доменов), они могут «включать» или «выключать» их активность (имеются в виду ферментативная активность, ДНК-связывающая способность и способность образовывать белок-белковые комплексы).
Центростремительная волна митогенной импульсации в максимально упрощенном виде сводится к передаче фосфатной группы наподобие эстафетной палочки от одной протеин-киназы к другой. В конечном итоге она достигает ядерных регуляторных белков, активирует их посредством фосфорилирования и тем самым индуцирует перепрограммирование генома. Необходимо отметить, что активность протеинкиназ практически на любом этапе переноса митогенного сигнала уравновешивается активностью противодействующих им ферментов —-дефосфорилирующих белки фосфатаз. Баланс позитивных и негативных эффектов — фундаментальное свойство регуляции клеточного деления, проявляемое на любом его уровне.
Противоположно направленный (центробежный, из ядра в цитоплазму) поток информации в виде молекул мРНК обусловливает специфическую реакцию клетки на митогенный сигнал — синтезируется множество новых белков, выполняющих структурные, ферментативные и регуляторные функции.
В структуре многих сигнальных белков существуют своеобразные «стыковочные узлы» разных типов, предназначенные для белок-белковых взаимодействий. Поскольку одна молекула может обладать несколькими такими участками, то существует возможность самосборки очень сложных многокомпонентных конструкций, необходимых для переноса сигнала и для регуляции транскрипции. Присоединение к конструкции новых элементов иногда обозначают термином «рекрутирование». Одни и те же структурные блоки могут формировать существенно разные конструкции, что наделяет систему функциональной гибкостью и свойством взаимозаменяемости отдельных ее элементов.
Нерегулируемое размножение трансформированной клетки можно представить, если продолжить аналогию с рефлекторной дугой, как следствие возникновения очага «застойного» возбуждения в том или ином звене пути переноса митогенного сигнала. Повреждение гена и, как следствие, структурный дефект какого-либо из сигнальных белков, способный зафиксировать его в постоянно активном состоянии (т.е. сделать независимым от «вышестоящих» регуляторных инстанций), — один из главных механизмов канцерогенеза.
Нормальные гены, участвующие в переносе митогенного сигнала и потенциально способные на такое превращение, называются протоонкогенами.
Баланс позитивных и негативных факторов, как уже отмечалось выше, — фундаментальное свойство любой сложной регуляторной системы, в том числе и управляющей клеточным делением. Протоонкогены — элементы позитивной регуляции; они являются акселераторами клеточного деления и в случае превращения в онкогены проявляют себя как доминантный признак.
Вместе с тем в давних опытах по образованию гетерокарионов (продуктов слияния клеток в культуре) установлено, что свойство туморогенности (способности образовывать опухоли при перевивке животным) ведет себя как признак рецессивный — гетерокарионы. образованные слиянием нормальных и трансформированных (опухолевых) клеток, ведут себя как нормальные. Таким образом, в нормальных клетках явно присутствуют факторы, тормозящие клеточное деление и способные при внесении в опухолевую клетку нормализовать ее. Многие из этих белковых факторов идентифицированы; кодирующие их гены получили название генов-супрессоров.
{kind=link}
Итак, полная трансформация клетки является следствием нескольких генетических событий — активации онкогена(ов) и инактивации гена(ов), осуществляющих супрессорные функции.
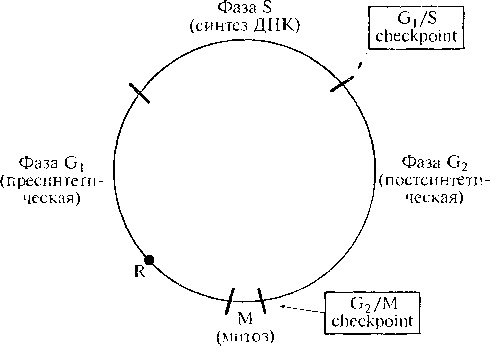
В основе канцерогенеза лежит нарушение цикла деления. Клетки организма находятся в одном из трех возможных состояний (рис. 12.3):
- в цикле;
- в стадии покоя с сохранением возможности вернуться в цикл;
- в стадии окончательной дифференцировки, при которой способность делиться полностью утрачена (таковы, например, нейроны головного мозга). Образовывать опухоли могут, естественно, только клетки, способные делиться.
Цикл удвоения разных клеток человека существенно варьирует: от 18 ч у клеток костного мозга до 50 ч у клеток крипт толстой кишки. Основными его периодами являются митоз (М) и синтез ДНК (фаза S), между которыми выделяют два промежуточных периода — G, и G2. Вовремя интерфазы (период между двумя делениями) клетка растет и готовится к митозу.
На протяжении фазы G1 существует ответственный момент (так называемая точка рестрикции R), когда решается, войдет ли клетка в следующий цикл деления или предпочтет стадию покоя G0, в которой она может находиться неопределенно долго. Как уже упоминалось, окончательно дифференцированные клетки постоянно находятся в стадии покоя, тогда как сохранившие способность к делению могут вернуться в цикл при соответствующей стимуляции внешними факторами, причем последующие этапы совершаются автоматически.
В отличие от «асоциальной» опухолевой клетки нормальная клетка подчиняется исходящим из организма сигналам (митогенный стимул). Если в определенный момент у нормальной клетки есть необходимые условия (достаточная масса и содержание белков, концентрация кальция, обеспеченность питательными веществами) и она к тому же получает митогенный стимул, то она вступает в очередной цикл деления. В отсутствие внешнего сигнала нормальная клетка выходит из цикла и в этом заключается ее коренное отличие от клетки опухолевой, которая побуждается к делению эндогенными стимулами.
При делении клетки есть два критически важных момента: фаза синтеза ДНК и вхождение в митоз, когда действуют своеобразные «контрольно-пропускные пункты» (checkpoints). В этих «пунктах» проверяется готовность к удвоению (репликации) ДНК (в первом случае) и завершенность репликации (во втором случае). Если в клетке ДНК повреждена, то ее удвоение блокируется перед началом деления. Следовательно, блокируется этап, способный закрепить повреждения ДНК и передать их потомству. Аналогичная цель достигается апоптозом, причем какой путь выберет клетка (блокировку деления или апоптоз) зависит от многих условий.
Процесс репликации ДНК продолжается несколько часов. За это время весь генетический материал должен быть воспроизведен абсолютно точно. В случае каких-либо отклонений продвижение клетки в цикле блокируется или она может подвергнуться апоптозу. Если же дефектны сами «контрольно-пропускные пункты», то дефекты генома не устраняются, передаются потомству и возникает опасность злокачественной трансформации клетки.
Как говорилось ранее, для деления клетки необходим митогенный сигнал, перенос которого — процесс многоэтапный.
В зависимости от типа клетки и конкретного митогенного стимула реализуется один из множества сигнальных путей. Например, ростовые факторы действуют путем, опосредованным тирозинпротеинкиназными рецепторами и MAP (mitogen activated protein) — киназным каскадом, т.е. каскадом реакций фосфорирования, возникающим как следствие митогенной активации клетки.
Ростовые факторы (регуляторы пролиферации).
Ростовые факторы секретируются одними клетками и действуют паракринным образом на других. Это небольшие белки; полипептидная цепь EGF (epidermal growth factor) состоит, например, из 53 аминокислот. Существует несколько семейств ростовых факторов, члены каждого из которых объединены структурной гомологией и функциональным сходством. Одни из них стимулируют пролиферацию (например, EGF и PDGF — platelet-derived growth factor, тромбоцитарный фактор роста), а другие (TGF-p, TNF, интерфероны) — подавляют ее.
Рецепторы к ростовым факторам.
Рецепторы расположены на клеточной поверхности. Каждая клетка обладает своим особым набором рецепторов и соответственно — набором ответных реакций. Тирозинкиназные рецепторы состоят из нескольких доменов: внеклеточного (взаимодействующего с лигандом), трансмембранного и подмембранного, обладающего тирозин-протеинкиназной активностью.
При связывании с ростовыми факторами (например EGF) молекулы рецепторов инициируют реакции, вследствие чего возникает трансмембранный перенос сигнала — зарождение той волны «возбуждения», которая распространяется затем в виде каскада реакций фосфорилирования внутрь клетки и благодаря которой митогенный стимул достигает в конце концов генетического аппарата ядра.
Ras-белки..
Одним из наиболее важных является сигнальный путь с участием Ras-белков (это подсемейство так называемых G-белков, образующих комплексы с гуаниловыми нуклеотидами; Ras-GTP — активная форма, Ras-GDP — неактивная). Этот путь один из основных в регуляции клеточного деления у высших эукариот — настолько консервативен, что его компоненты способны заменить соответствующие гомологи в клетках дрозофилы, дрожжей и нематод. Он опосредует разнообразные сигналы, исходящие из внешней среды, и функционирует, по всей вероятности, в каждой клетке организма. Ras-белки играют роль своеобразного турникета, через который должен пройти почти любой из поступающих в клетку сигналов. Критическая роль этого белка в регуляции клеточного деления известна с середины 80-х г. XX в., когда активированная форма соответствующего гена (онкоген Ras) была обнаружена во многих опухолях человека.
Ras-опосредованный сигнальный путь контролирует так называемый МАР-киназный каскад. Активность ферментов, участвующих в киназных каскадах, уравновешивается активностью противодействующих им и находящихся под столь же строгим контролем фосфатаз. Результатом активации МАР-киназ является индукция ряда факторов транскрипции и, как следствие, стимуляция активности ряда генов.
Таким образом, если в нормальной клетке активная конформация сигнального белка формируется лишь под воздействием внешнего стимула и имеет транзиторный характер, то в клетке трансформированной (и ее клонах) она закреплена постоянно.
Активация транскрипции.
Группа генов, определяющих вхождение клетки в фазу S, активируется транскрипционным фактором АР-1, представляющим собой комплекс белков — членов семейств Jun и Fos (гены, их кодирующие, — c-Jun и c-Fos — относятся к числу протоонкогенов; буква «с» (от англ. cell) обозначает их клеточное происхождение в отличие от вирусных онкогенов v-Jun и v-Fos). Эти транскрипционные факторы могут взаимодействовать между собой с образованием множества гомо- и гетеродимеров, связывающихся с определенными участками ДНК и стимулирующих синтез РНК на прилежащих к этим участкам генов.
МАР-киназы повышают активность АР-1 двумя способами:
- опосредованным, активируя соответствующие гены;
- прямым, фосфорилируя входящие в их состав сери новые и треониновые остатки.
В результате активации генов продуцируются белки, необходимые для синтеза ДНК и последующего митоза. Некоторые из новообразованных белков, известные как белки раннего ответа, выполняют регуляторные функции: связываясь со специфическими участками ДНК, они активируют прилежащие гены. Другую группу белков составляют ферменты, такие как тимидинкиназа, рибонуклеотидредуктаза, дигидрофолатредуктаза, тимиди-латсинтетаза, орнитиндекарбоксилаза, ДНК-полимеразы, топоизомеразы и множество других, которые имеют непосредственное отношение к синтезу ДНК. Усиливается, кроме того, общий белковый синтез, поскольку при каждом цикте удвоения воспроизводятся все клеточные структуры.
Нормальное прохождение клеткой отдельных фаз клеточного цикта обеспечивается строгой последовательностью и упорядоченностью многочисленных ферментативных процессов. Роль «водителя ритма» при этом играют циклины — белки, закономерным образом сменяющие друг друга на протяжении цикла.
Циклические реакции.
Реализация митогенного сигнала происходит следующим образом. Имеется два семейства белков, «движущих» клеточный цикл — циктин(о>с/ш)-зависимые серин/треониновые протеинкиназы и сами циклины. Последние синтезируются и распадаются в строго определенные моменты цикта, различные для разных циклинов. Имеются три основных их класса: G1-циклины, необходимые для прохождения G,/S, S-циклины — для прохождения S-фазы и G2 (митотические) циклины — для вхождения в митоз. Исчезновение циклина из внутриклеточной среды в определенный момент времени столь же важно, как и его появление. Например, в митозе (на границе метафазы и анафазы) один из циклинов быстро деградирует в результате протеолиза, если этого не происходит, то митоз не может завершиться и разделения дочерних клеток не происходит.
Контрольно-пропускные пункты.
Любое стрессорное воздействие (например, отсутствие питательных веществ, гипоксия и особенно повреждение ДНК) блокирует движение клетки по циклу в одном из двух контрольных пунктов (checkpoints). Во время этих остановок активируются механизмы надзора, способные: обнаружить повреждение ДНК; передать сигнал неблагополучия, блокирующий синтез ДНК или митоз; активировать механизмы репарации ДНК. Благодаря этому обеспечивается стабильность генома. Механизм контроля G1/S блокирует репликацию ДНК и активирует процессы репарации (или индуцирует апоптоз), тогда как до завершения репликации митоз запрещен. Дефекты этих механизмов могут привести к появлению дочерних клеток с поврежденным геномом.
В механизме checkpoint участвуют комплексы Cdk-cyclin и ряд дополнительных белков — КЬ, р53 и др. Их совокупность образует систему «тормозов», не позволяющих клетке делиться в отсутствие адекватных стимулов.
Кодирующие эти белки гены получили название генов-супрессоров. Особое значение этой системы заключается в том, что раковая трансформация клетки возможна только после инактивации генов-супрессоров. Важно при этом иметь в виду, что в соматической клетке существуют по два аллеля каждого из генов, в том числе и генов-супрессоров, и. следовательно, для их инактивации необходимы два независимых события (например, деления одного аллеля и мутация другого). Именно в этом кроется причина того, что «спорадические» опухоли возникают относительно редко (вероятность возникновения в одной клетке нескольких независимых мутаций, причем поражающих один и тот же локус обеих хромосом, относительно невелика), а «семейные» чрезвычайно часты (в «раковых» семействах один из двух наследуемых аллелей того или иного гена-супрессора исходно дефектен). В последнем случае система «тормозов» у всех клеток данного организма держится лишь на одном нормальном аллеле, что резко снижает ее надежность и повышает риск возникновения опухоли. Именно это и происходит при наследственной ретинобластоме и других наследственных заболеваниях.
В механизме контроля фазы клеточного цикла G1/S участвуют ингибирующие белки, в частности семейство СК I (Cdk inhibitory proteins), состоящее из нескольких членов (р27, рl6, р21), которые продуцируются клеткой в ответ на разные стимулы и блокируют клеточное деление. Таким стимулом является, например, TGFb (transforming gowth factor b), который активирует белок р27. И наоборот, митоген интерлейкин-2 (ИЛ-2) инактивирует р27 и способствует тем самым продвижению клеток в цикле деления. Этот же белок р27 участвует, по-видимому, в феномене контактного ингибирования (блокировке деления клеток при соприкосновении их поверхностей) — характерном свойстве нормальных клеток, отсутствующем у опухолевых клеток.
Сходную роль ингибитора клеточного деления выполняет СК 1 рl6, который кодируется геном-супрессором. СК I р16 инактивирован во многих линиях опухолевых клеток и в клетках «семейной» меланомы.
Особо важную роль в формировании блока G1/S играет белок р21. Его синтез индуцируется главным клеточным «хранителем генома» — белком-супрессором р53. В свою очередь р53 активируется при возникновении различных повреждений ДИК и, в частности, при гамма-облучении клеток.
В клетках с интактной ДНК содержание р53 исчезающе мало и соответственно ничто не препятствует активации группы генов, необходимых для вступления клетки в фазу синтеза ДНК. У клеток с дефектным или отсутствующим геном р53 контрольный пункт G1/S неполноценен. Это проявляется в том, что повреждения ДНК, индуцированные ионизирующей радиацией или каким-либо другим способом, не приводят ни к задержке клеток на границе фаз G1/S, ни к апоптозу. В результате в популяции накапливаются клетки со множественными нарушениями структуры ДНК; появляется и со временем нарастает нестабильность генома, которая ведет к появлению все новых клонов клеток. Их естественный отбор лежит в основе опухолевой прогрессии — постоянного «дрейфа» опухоли к все увеличивающейся автономности и злокачественности. Именно свойство нестабильности генома наделяет клетки опухоли, с одной стороны, замечательной приспособляемостью к условиям окружающей среды и «увертливостью» по отношению к лечебным воздействиям, а с другой — все новыми качествами, в частности, способностью метастазировать.
«Выше» р53 в регуляторной цепи расположен белок, кодируемый геном А73 (ataxia-telangiectasia mutated). Его функция состоит в активации р53 в ответ на повреждение ДНК ионизирующей радиацией. В свою очередь р53 активирует группу генов, продукты которых, по-видимому, «запускают» механизм репарации.
Апоптоз
Апоптоз (программируемая клеточная гибель) — широко распространенный биологический феномен клеточного самоуничтожения, которое индуцируется либо разнообразными внешними стимулами, либо внутренними условиями клетки, невозможностью репарации повреждений ДНК.
Роль апоптоза велика не только в формообразовательных процессах во время эмбриогенеза (формирование органов, замена одних тканей другими, резорбция временных органов и т.д.), но и в поддержании тканевого гомеостаза во взрослом организме.
У опухолевых клеток программа клеточной гибели во многих случаях блокирована, что вносит существенный вклад в увеличение массы опухоли.
Существует два типа клеточной гибели: некроз и апоптоз (клеточное «убийство» и «самоубийство», соответственно), различающиеся как индуцирующими эти процессы стимулами, так и механизмом их осуществления.
Некроз — это метаболическая катастрофа, вызванная тяжелыми повреждениями клеточных структур. Для него характерно раннее увеличение объема клетки и митохондрий с последующим их аутолизом, тогда как внутриядерные изменения обнаруживаются позднее.
Апоптоз как явление физиологическое распространен гораздо шире. С его участием проходят морфообразовательные процессы в эмбриогенезе, позитивная и негативная селекция Т- и В-лимфоцитов, индуцированная глюкокортикоидами гибель лимфоцитов, а также гибель клеток при юс естественном старении. Усиленный апоптоз ведет к инволюции органа, что встречается и в физиологических условиях (например, молочная железа в постлактационном периоде) и при патологии (например, при дегенеративных заболеваниях центральной нервной системы).
Опухолевые клетки имеют селективное преимущество над нормальными, поскольку механизм апоптоза у них зачастую дефектен. Кроме того, у многих из них развивается резистентность к различным химиопрепаратам и к облучению, которые вызывают гибель также по механизму апоптоза. Морфологически апоптоз проявляется вначале в конденсации хроматина, уменьшении объема клетки и фрагментации клеточного ядра, в характерной складчатости плазматических мембран.
Одним из ключевых моментов апоптоза является распад ДНК. Процесс заканчивается фрагментацией клетки с образованием так называемых апоптотических телец, которые быстро фагоцитируются макрофагами или соседними клетками без сопутствующей воспалительной реакции.
Белки, участвующие в апоптозе, подразделяют на две группы в соответствии с их антиапоптическим или проапоптическим действием. Комплексирование проапоптотических и антиапоптических белков ведет, по-видимому, к их взаимному «погашению», поэтому результат (индукция апоптоза или, напротив, его отмена) зависит от «баланса сил». Гибель опухолевых клеток под воздействием химио-препаратов и облучения происходит по механизму апоптоза.
Механизмы опухолевой трансформации
Принципиальное различие между нормальной и опухолевой клетками заключается в том, что переход от покоя к делению (G0 —> G1) в первом случае инициируется внешними по отношению к клетке стимулами, а во втором — внутренними. Молекулярные механизмы этого явления сводятся к активации онкогенов и инактивации генов-супрессоров в опухолевых клетках.
Активация онкогенов.
В середине 1980-х гг. было доказано, что некоторые нормальные клеточные гены обладают онкогенным потенциалом. Так сформировалась концепция онкогена, возникающего из нормального гена (протоонкогена) при его повреждении тем или иным способом. Такая трансформация называется активацией онкогена. Протоонкогены — «акселераторы» клеточного деления. Их функция проявляется как доминантный признак, т.е. становится явной при активации даже одного аллеля (выпадение тормозящей функции генов-супрессоров обнаруживается лишь при утере обоих аллелей).
Возникает вопрос, какие именно гены могут трансформироваться из прогоонкогенов в онкогены? Установлено, что этим свойством обладают гены, участвующие позитивным образом в переносе митогенного сигнала независимо от занимаемого ими в сигнальном пути места (начиная с факторов роста и заканчивая транскрипционными факторами). Число известных протоонкогенов достигает сегодня нескольких десятков и постоянно растет по мере углубления представлений о механизмах клеточного деления. При повреждении какого-либо из этих генов тем или иным способом (факторами физическими или химическими, эндогенными или экзогенными) структура кодируемого им белка может оказаться жестко зафиксированной в процессе активной конформации. Таким образом, в нормальной клетке воздействие внешнего стимула вызывает активную конформацию сигнального белка, которая продолжается лишь до тех пор, пока действует этот стимул.
В мутантной клетке (и в ее клонах) активная конформация белка оказывается жестко закрепленной. Ущербный сигнальный белок «гонит волну возбуждения» вниз по течению (downstream) независимо от состояния вышестоящих структур. Возникает состояние «застойного возбуждения», которое лежит в основе автономного, нерегулируемого деления опухолевой клетки. Приведем несколько примеров. В разных «спонтанных» опухолях человека (рак желудка, мочевого пузыря и т.д.) обнаружена точковая мутация с-Ha-ras-l (одного из представителей семейства ras). Оказалось, что достаточно замены одного основания (соответственно одной аминокислоты в белке), чтобы протоонкоген превратился в онкоген, т.е. приобрел трансформирующие свойства. При этом кодируемый геном c-Ha-ras-1-белок теряет в определенной мере ПФазную активность, поэтому находится постоянно в «возбужденном» состоянии, т.е. в отличие от нормальных клеток, где митогенный сигнал имеет импульсный характер, в клетках с поврежденным ras митогенная стимуляция непрерывна. Такое самоподдерживающееся возбуждение выводит клетку из-под регуляторных влияний организма.
Активация онкогенов может происходить двумя путями — в результате дефекта регуляции их транскрипции или изменения «дозы» гена (при его амплификации), что приводит к избыточному синтезу соответствующего белка, не адекватному месту и времени.
Таким образом, активация онкогена обусловлена нарушением его регуляторной или структурной части.
Инактивация генов-супрессоров.
Как уже упоминалось, для полной трансформации клетки в подавляющем большинстве случаев недостаточно одной лишь активации онкогена. Бесконтрольному клеточному размножению препятствуют гены-супрессоры. Об их существовании догадывались давно, исходя из двух групп фактов. Во-первых, в опытах по слиянию клеток и образованию гетерокарионов было установлено, что в паре «клетка нормальная + клетка злокачественная» «берет верх» первая (полученный соматический гибрид теряет свои туморогенные свойства). Это свидетельствовало о наличии в нормальной клетке некого тормозящего митотическую активность фактора, отсутствующего, по-видимому, в опухолевой клетке, и о том. что соответствующие гены рецессивны — клетки становятся опухолевыми при утере обоих аллелей. Во-вторых, при цитогенетическом исследовании опухолевых клеток постоянно обнаруживаются делении хромосом, довольно специфические для разных форм рака. Это заставляло думать (и предположение полностью оправдалось), что именно отсутствие соответствующих генов лишает клетку митотических «тормозов».
Например, цитогенетический анализ больных ретинобластомой (злокачественная опухоль сетчатки глаза, характеризующаяся зачастую наследственной предрасположенностью) выявляет характерную для этой формы делению в 13-й хромосоме, где при последующем анализе действительно был идентифицирован ген-супрессор Rb. Наследственная делеция одного аллеля этого гена делает человека предрасположенным к заболеванию ретинобластомой или остеосаркомами.
Гены-супрессоры кодируют белки, блокирующие, каждый своим особым образом, разные этапы митогенной стимуляции (к ним относятся уже упоминавшиеся р53 — главный «хранитель генома», Rb — формирующий механизм «G1/S checkpoint», p16 и р21, блокирующие активность комплексов cyclin-Cdk, и др.).
Ген-супрессор р53. Особую роль в канцерогенезе играет ген-супрессор р53. Продукт этого гена (белок с молекулярной массой 53 кДа) — регулятор транскрипции, способный активировать одни гены и подавлять другие, дефектен в большинстве опухолей человека, независимо от их происхождения и локализации (опухоли легких, головного мозга, толстой кишки, пищевода, молочной железы, шейки матки, печени, гемопоэтических и ретикулоэндотелиальных тканей и др.). Белок р53 чрезвычайно консервативен (обнаружен у многих представителей животного мира), состоит (у человека) из 393 аминокислот, локализован, как правило, в клеточном ядре.
Функция р53 в нормальных клетках заключается в связывании со специфическими последовательностями ДНК, находящимися в промоторных участках других генов, и в управлении их активностью. Повреждение ДНК нормальных клеток приводит к увеличению содержания р53 и к индукции р53-зависимых белков. Это в свою очередь вызывает задержку клеток на границе G1/S или к апоптозу.
Во многих опухолевых клетках (возможно, во всех) гены р53 делецированы или мутантны. Мутации по-разному в зависимости от локализации инактивируют р53. Возможны и другие причины функциональной инактивации р53. Например, в результате инфицирования клеток некоторыми вирусами, продукты которых способны взаимодействовать с р53 (такова, по-видимому, одна из причин возникновения рака шейки матки у женщин, инфицированных вирусом папилломатоза человека HPV). К тому же результату приводит продукция в избыточном количестве некоторых внутриклеточных белков, способных связывать р53 и лишать его функциональной активности.
Клетки с дефектами р53 имеют ряд существенных особенностей — резко возрастающую нестабильность генома и дефекты контрольного пункта G1/S. Таким образом, ключевая роль р53 заключается, по всей вероятности, в поддержании стабильности генома и в индукции апоптоза в случае нерепарируемых повреждений ДНК run неразрешимых внутриклеточных конфликтов. Функциональная инактивация р53 способствует трансформации нормальных клеток в опухолевые, с одной стороны, и прогрессии уже возникшей опухоли — с другой.
{kind=link}
Белок р53 участвует в опухолевой прогрессии и совершенно особым образом. Оказалось, что низкое содержание кислорода в слабо васкуляризированных участках ткани стимулирует синтез р53 и вследствие этого происходит апоптоз (рис. 12.4). Мутации р53 в опухолевой клетке отменяют, однако, апоптоз. В результате опухолевые клетки с инактивированным р53 получают селективное преимущество над интактными. Таким образом, именно гипоксическая среда является, возможно, тем плавильным котлом. из которого появляются р53-мутанты.
«Как в сцене из дантова «Ада» рождаются эти уродливые клетки среди распадающихся останков своих предков, неуязвимые и благополучные в условиях, совершенно пагубных для простых смертных… Глухие к нормальным клеточным регуляторам, они бесконтрольно размножаются и постоянно увеличивают степень своей ненормальности благодаря амплификации генов и хромосомным перестройкам. Это только вопрос времени, когда они выберутся из своего обиталища, проникнут в окружающие нормальные ткани и нападут на своих невинных соседей…» (К. Кинцлер, Б.Вогельштейн; K.KinzIer, В. Vogelstein, 1996).
Ген-супрессор ЛРС (Adenomatous Polyposis Coli).
Этот ген-супрессор часто дефектен при синдромах семейного аденоматозного поли поза и наследственного неполипозного рака толстой и прямой кишки. В толстом кишечнике пациентов с наследственными мутациями АРС во II —III декаде жизни возникают тысячи аденоматозных полипов, часть из которых могут дать начало опухоли (вторая по значимости причина онкологической смертности в США).
АРС — первый из белков-супрессоров, которые наделяют особой функцией «сторожа» (gatekeeper), обеспечивающего тканевый гомеостаз.
Эти белки ответственны за постоянство числа клеток в постоянно обновляемых тканях и за адекватность клеточных реакций в ситуациях, требующих роста ткани (например, при ее развитии или травме). Повреждение «сторожа» приводит к нарушению тканевого гомеостаза из-за дисбаланса клеточной гибели и пролиферации. Ген с подобной функцией находится в уникальном положении — его нарушение инициирует пролиферацию и, если последуют мутации других генов, рост опухоли. Напротив, повреждения иных генов при нормальном «стороже» не могут иметь никаких последствий. Отсюда следует, что определяющим для канцерогенеза может оказаться не простое накопление мутаций в тех или иных генах, а порядок появления этих мутаций. Если уже первая из них затрагивает АРС, 70 последующие канцерогенные события весьма вероятны, поскольку дефектные клетки размножаются. увеличивая «базу» для последующих мутационных событий. Напротив, если первичные события затрагивают другие гены, то функционально активный «сторож» срабатывает, не позволяя этим клеткам делиться и индуцируя их апоптоз.
Гипотеза «сторожа» возникла как результат анализа мутационного процесса при колоректальном раке человека и животных, выявившего повреждения АРС на самых ранних стадиях канцерогенеза (уже на стадии микроскопических узелков), тогда как мутации других генов (в частности, RASnpSS) возникают значительно позже. Кроме того, мутации р53 обнаруживают в клетках тканей, видимым образом не измененных. Это дает основание для предположения, что онкогенный потенциал мутаций может проявиться только на фоне дефектного АРС, который играет определяющую роль в инициации рака, тогда как дефекты других генных продуктов (онкогенов и супрессоров) — в его прогрессии. Предполагается, что роль «сторожа» в разных тканях исполняют разные гены.
Нарушения механизмов репарации Днк.
Инициация и прогрессия опухоли — это фенотипические проявления повреждений генотипа (ДНК). Повреждения ДНК различны при химическом мутагенезе, ультрафиолетовом облучении, при воздействии ионизирующей радиации. Соответственно этому существует и несколько типов репарации, которые выполняют очень важную защитную функцию. При некоторых наследственных синдромах (пигментная ксеродерма, синдром Линча и др.) механизмы репарации неполноценны. В результате у таких больных развиваются множественные опухоли разных тканей.
Таким образом, в области экспериментальной онкологии сделаны фундаментальные открытия, значительно продвинувшие нас в понимании не только того, как нормальная клетка трансформируется в опухолевую, но и как она функционирует, рождается и погибает.
Менее существенны достижения в лечении злокачественных новообразований. Не удалось пока выявить такие биохимические особенности опухолевых клеток, которые позволяли бы, с одной стороны, идентифицировать процесс на самых ранних, еще доступных лечению стадиях его развития, и, с другой стороны, найти ту «магическую пулю», которая избирательно поражала бы злокачественные клетки, не затрагивая нормальные. Вместе с тем есть основания для осторожного оптимизма. Во-первых, становятся все более эффективными профилактические мероприятия, способные резко снизить онкологическую заболеваемость (профилактика курения, алкоголизма, других вредных привычек, экологические мероприятия, улучшение условий труда). Наряду с классической триадой (хирургия, химиотерапия, облучение) для лечения онкологических больных начинают применяться принципиально новые методы — иммунотерапия и фотодинамическая терапия, а также биотехнологические подходы, направленные на активацию в опухолевых клетках генов-супрессоров и проапоптических генов. Весьма обнадеживающие результаты получены в поиске препаратов, блокирующих формирование сосудистого ложа в опухолевой ткани. Принимая во внимание быстрый прогресс во всех этих направлениях, можно ожидать ощутимых практических результатов уже в ближайшем будущем.
Контрольные вопросы
- Что такое опухоль?
- Чем отличается доброкачественная опухоль от злокачественной?
- Какие факторы способствуют образованию опухоли?
- Какие этиологические факторы канцерогенеза вам известны?
- Какие существуют стадии развития опухоли?
- Какие свойства отличают опухолевые клетки от нормальных?
- В чем выражается системное действие опухоли на организм?